
Онтогенезом называют индивидуальное развитие организма от зиготы (или вегетативного зачатка) до естественной смерти. В ходе онтогенеза реализуется наследственная информация организма в конкретных условиях окружающей среды, в результате чего формируется совокупность всех признаков и свойств данного индивидуального организма.
Каждый растительный организм в своем развитии проходит ряд этапов, характеризующихся морфологическими и физиологическими особенностями. Развитие высших растений подразделяют на четыре этапа: 1) эмбриональный этап, 2) ювенильный (молодость), 3) репродуктивный (зрелость), 4) старость.
I. Этапы онтогенеза высших растений
1. Эмбриональный этап
Эмбриональный этап онтогенеза растения семенных растений охватывает развитие зародыша от зиготы до созревания семени включительно.
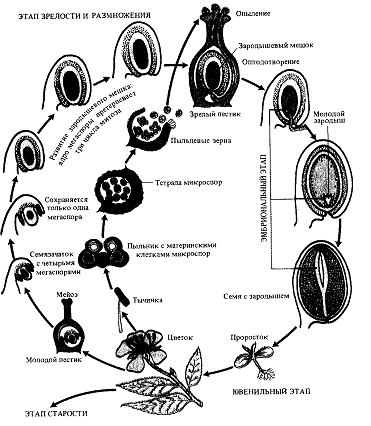
Рисунок 1. Этапы онтогенеза и фазы развития покрытосеменных растений
Зигота у покрытосеменных образуется в результате слияния спермия пыльцевой трубки (мужской гаметофит) с яйцеклеткой зародышевого мешка (женский гаметофит).
В зародышевом мешке у покрытосеменных происходит двойное оплодотворение, состоящее в слиянии второго спермия пыльцевой трубки со вторичным (диплоидным) ядром центральной клетки зародышевого мешка, что в дальнейшем приводит к образованию эндосперма. Зародышевый мешок находится в нуцеллусе, окруженном интегументом. Нуцеллус используется для питания зародыша и реже превращается в запасающую ткань — перисперм.
Яйцеклетка в зародышевом мешке уже поляризована, т. е. две ее половины, обращенные к микропиле (пыльцевходу) и к халазе (основанию), имеют разное строение: ядро сдвинуто к халазальному полюсу, а в микропилярной половине расположена крупная вакуоль.
Рисунок 2. Строение семяпочки ириса (по Додель-Порт)
Зародыши проходят ряд последовательных фаз развития. Для большинства двудольных это проэмбрио, глобулярная, сердцевидная, торпедовидная (торпедо) и созревания.
После оплодотворения зигота некоторое время находится в латентном состоянии. За это время в ней возрастает синтез РНК, она увеличивается в объеме. Триплоидное ядро центральной клетки начинает делиться первым, образуя эндосперм.для развития эндосперма нужны ИУК и цитокинин, которые поступают из нуцеллуса и из плаценты.
При первом делении зиготы, плоскость которого перпендикулярна к оси её поляризации, дочерняя клетка, обращенная у микропиле, выглядит более крупной и при дальнейшем делении образует однорядную нить клеток, называемую суспензором. Функции суспензора: 1) удлиняясь, вдвигать развивающийся зародыш в ткань эндосперма, 2) служить гаусторией, поглощающей вещества из нуцеллуса и интегумента и передающей их зародышу, 3) синтезировать фитогормоны. Часть клеток суспензора, примыкающая к зародышу, у некоторых видов входит в его состав в качестве апикальной меристемы и корневого чехлика зародышевого корня. Две синергиды, примыкающие к яйцеклетке, выполняют функцию гаусторий, поглощая вещества из нуцеллуса. Аналогичную функцию гаусторий осуществляют и клетки-антиподы, находящиеся рядом с эндоспермом.
Новая методология периодизации онтогенеза растений
... На рис. 4. иллюстрируется жизненный цикл растения сои. Исследования развития растений кукурузы показало, что у среднеспелого гибрида Моника продолжительность онтогенеза составляет 411 дней, а вегетационного периода ... цветка, микро - и макроспорогенез в закрытом бутоне. 9 – формирование пыльцы и зародышевого мешка, микро - и макрогаметогенез. Интенсивный рост верхних междоузлий стебля. 2. Ювенильный ...
Вторая клетка у двухклеточного зародыша обращенная к эндосперму и халазальному полюсу, дважды делится в продольном по отношению к первичной оси поляризации направлении, образуя квадрант. Затем каждая из четерех клеток будущего зародыша у многих двудольных делится поперек, в результате чего формируется октант. Четыре дистальные клетки октанта при дальнейшем делении сформируют семядоли и апекс побега, а четыре проксимальные — гипокотиль и базальную часть корня. На этой фазе развития зародыша еще нет видимой дифференциации клеток, и зародыш, состоящий последовательно из одной, двух, четырех и восьми клеток, называют проэмбрио. По мере развития зародыша концентрация ИУК в клетках постепенно возрастает, происходит постоянный приток цитокинина. Последняя клетка суспензора, примыкающая к октанту, становится гипофизой — инициальной клеткой корневого полюса и морфологически выделяется.
Все восемь клеток проэмбрио делятся периклинально, из-за распределения сил механического давления, направленного изнутри к поверхности. Эта фаза развития называется глобулярной. Наружные клетки в дальнейшем сформируют протодерму, а внутренние дадут начало первичной каре и центральному цилиндру. На этой фазе особенно необходим цитокинин. Фитогормоны поступают главным образом из развивающегося эндосперма, который играет ведущую роль в развитии зародыша.
Следующая фаза развития зародыша получила название сердцевидной. В морфологически верхней части глобулярного зародыша наблюдается интенсивное билатеральное деление клеток, в результате чего закладываются примордии двух симметрично расположенных семядолей. В участке между ними (будущий апекс побега) деление клеток, наоборот, резко замедляется. В этот период для нормального развития необходим приток ИУК, цитокинина и аденина.
Торпедовидная фаза развития зародыша связана с делением клеток преимущественно поперек продольной оси и с более интенсивным ростом клеток в зачатках семядолей и в зоне гипокотиля. Отчетливее выделяются по вытянутой в длину форме клетки прокамбия и гипокотиле. Формируются промеристема корня. На этой фазе, кроме всё более возрастающей концентрации ИУК и присутствия цитокинина, необходим гиббереллин (для роста гипокатиля).
У некоторых двудольных удлиняющиеся семя доли и гипокатиль изгибаются и складываются вдвое. Между семядолями закладывается апекс побега, происходит распад суспензера. Если семядоли выполняют функцию вместилища запасных веществ, то они заполняют почти весь объём созревающего семени, в них на последних этапах формирования зародыша откладываются запасные белки, крахмал и жиры.
Приток питательных веществ в развивающиеся семязачатки и затем в созревающие семена и формирующиеся плоды определяется тем, что эти участки становятся доминирующими центрами: в их тканях вырабатывается большое количество фитогормонов, прежде всего ауксина, в результате чего аттрагирующее действие этих тканей резко возрастает.
Развитие нервной системы
... результате деления клеток-предшественников вентрикулярной зоны нервной трубки. Постмитотические нейроны мигрируют от вентрикулярной поверхности нервной трубки и образуют серое вещество НС взрослых позвоночных. В пределах каждой из областей в развивающейся нервной системе ... открытие механизмов и молекул, которые управляют развитием нервной системы удивительно сходным образом у различных животных. ...
На последнем этапе созревания семени теряют значительное количество воды и у большинства видов средней полосы переходят в состояние покоя. Этот переход связан с уменьшением в тканях свободных ауксинов, цитокининов и гиббереллинов и с увеличением содержания АБК (Полевой, 1989).
Процессы роста, дифференциации и специализации растительных клеток сопровождаются значительными изменениями строения, состава и функциональных особенностей их оболочек. Эти изменения наиболее выражены при переходе к генеративному развитию, когда образуется гаплоидный гаметофит. Двухслойная оболочка мужского гаметофита состоит из экзины (наружный слой) и интины (внутренний слой).
Поровые области оболочки, через которые может прорасти пыльцевая трубка, отличаются редуцированной экзиной и утолщенной интиной. Экзина состоит, главным образом, из спорополленина, который представляет собой полимерную сеть, образованную из цепей небольших алифатических мономеров и остатков окисленных ароматических кислот. Интина включает три слоя: внутренний, средний и наружный. Начальный этап прорастания пыльцевого зерна подготавливает формирование и последующий быстрый рост пыльцевой трубки. Он начинается с гидратации пыльцевого зерна, обезвоженного в период созревания, и включает активацию метаболических и транспортных процессов, а также структурную реорганизацию цитоплазмы вегетативной клетки. В процессе гидратации, благодаря уникальной способности оболочки к растяжению, ее эластичности и прочности, менее чем за минуту достигается двукратное увеличение диаметра пыльцевого зерна. Одновременно вокруг пыльцевого зерна обнаруживаются диффундирующие из оболочки белки и пектины. Для соматических клеток и пыльцевой трубки показано, что пектины, транспортируемые в везикулах Гольджи, полностью этерифицированы, а в стенке они частично деметилируются под действием пектинметилэстераз (ПМЭ).
Добавление экзогенной ПМЭ увеличивает жесткость стенки трубки и ингибирует ее рост. При изучении пыльцевой трубки обнаружена прямая корреляция между степенью деэтерификации пектинов и жесткостью стенки по длине трубки. Таким образом, экспериментально установлена взаимосвязь между ростовыми процессами в клетке, физическими свойствами стенки и состоянием пектинов (Мейчик и др., 2009).
2. Ювенильный этап
Этап молодости у семенных растений начинается с прорастания семян или органов вегетативного размножения и характеризуется быстрым накоплением вегетативной массы. Растения в этот период не способны к половому размножению.
В ювенильный период осуществляется прорастание семян (или вегетативных зачатков) и формирование вегетативных органов. Прорастание в свою очередь делится на фазы набухания семян, проклевывания, гетеротрофного роста проростка, перехода к автотрофному способу питания.
По окончании у семян периода покоя поглощение ими воды служит пусковым фактором прорастания. Это поглощение осуществляется благодаря повышению в ряде случаев проницаемости семенных покровов для воды и за счет гидратации биополимеров в клетках. В результате развивается онкотическое давление (давление набухания) и семенные покровы разрываются. Набухание практически не зависит от температуры, содержания кислорода, освещения.
Гибридизация протопластов растений
... клубней занимали промежуточное положение между культурными и дикими растениями. Вместе с тем гибрид, полученный в результате соматической гибридизации, оказался устойчивым к вирусу "У", чем отличался ... хромосом одного из родителей. В культурах межсемейственных гибридов наблюдалось много многоядерных клеток, клеток с мини ядрами, в метафазах делений встречались гигантские хромосомы. Отмечена ...
Проклевывание начинается, когда семена достигают критической влажности (40 — 65% в пересчете на сырую массу), и происходит путем роста растяжением самого зародышевого корня или гипокотиля, в результате чего кончик корешка выталкивается из семени. Деление клеток наступает обычно позже. Рост растяжением клеток зародышевой оси обусловлен снижением содержания АБК при набухании семян. Выход корня обеспечивает закрепление прорастающего семени в почве и улучшает поглощение воды.
Вслед за корнем начинается рост побега. Прорастая в темноте (в почве), и корень, и побег ориентируются прежде всего по гравитационному вектору, при этом корень углубляется в почву, а побег — направляется к свету. Рост осевых частей зародыша проростка поддерживается фитогормонами. Причем у злаков ИУК и цитокинины вначале поступают в зародыш из эндосперма, ГА освобождается из связанного состояния в зародышевой оси, а через несколько часов индуцируется синтез гиббереллинов в щитке. Эпителиальные клетки щитка начинают переваривать запасные вещества в эндосперме. К процессу пищеварения подключаются клетки алейронового слоя, активированные гиббереллином. Подкисление тканей эндосперма за счет работы Н-помп создает условия для кислого пищеварения и для всасывания продуктов разложения крахмала и белков клетками эпителия. Клетки эпителия начинают вытягиваться и врастать в эндосперм, осуществляя гетеротрофный способ питания растущего зародыша и проростка.
Рост зародышевого корня сопровождается появлением вдоль него зон деления, растяжения и дифференциации клеток. Корень сам начинает синтезировать цитокинины и ГА, которые направляются в побег. Побег удлиняется благодаря растяжения гипокотиля (у бобов, тыквы и др.) или мезокотиля ( у злаков).
Листья не развиваются и гипокотиль в своей верхней части сильно изгибается наподобие крючка, что облегчает его перемещение в почве. В почечке у двудольных и в верхушке колеоптиля у злаков синтезируется ИУК.
Когда этиолированный побег достигает поверхности земли, возникает светоростовая и фотоморфогенетическая реакция: рост гипокотиля или мезокотиля резко подавляется, усиливается рост эпикотиля (первого настоящего междоузлия) и листьев. Содержание этилена в зоне крючка снижается и крючок распрямляется. Растение зеленеет и переходит к фототрофному типу питания.
Продолжительность ювенильного периода у разных видов растений неодинакова: от нескольких недель (однолетние травы) до десятков лет (у древесных).
Проростки по многим параметрам не похожи на взрослые растения. Различия часто хорошо заметны в форме листьев (хлопчатник, огурец, ипомея и др.).
У некоторых растений наблюдаются различия и во внутреннем строении. Так, у проростков папоротника проводящая система организована проще, чем у взрослых растений. У ювенильных растений менее мощная верхушечная меристема. Морфологическим признаком ювенильности в ряде случаев может служить также характер роста растения. Так, плющ в ювенильном состоянии представляет собой лазящий кустарник, а при переходе к взрослому состоянию образует вертикальный куст.
Старение как закономерный этап онтогенеза. Проявления старения ...
... жизни. Старение - заключительный этап онтогенеза. Старение - это закономерный процесс возрастных изменений организма, ведущей к изменению его адаптивной возможности и к увеличению вероятности смерти. Старение, закономерно ... клеток и сосудов, ацинарных клеток околоушной слюнной железы растет. Неодинаково изменяются и другие свойства клеток, например, прямая возбудимость скелетно-мышечных клеток ...
Этап молодости характеризуется полным отсутствием цветения или цветение слабо выражено даже при самых благоприятных для этого условиях. Здесь ярко проявляется роль компетенции. В данном случае ювенильное растение не обладает компетенцией к факторам, вызывающим закладку органов полового или вегетативного размножения. Это может быть связано с отсутствием в органах-мишенях, воспринимающих гормоны, белков-рецепторов, участвующих в индукции генеративного развития.
Однако отсутствие цветения само по себе не может служить показателем ювенильности, так как многие растения, находясь в зрелом состоянии, долго не зацветают без необходимых для этого условий. Поэтому более надежными критериями ювенильности служат описанные морфологические признаки.
Для ювенильных растений характерна значительно большая способность к корнеобразованию, что с давних пор используется в практике садоводства. Предполагается, что лучшая укореняемость ювенильных черенков — следствие более высокого содержания ауксинов. Однако обработка ауксином взрослых черенков не вызывает возврата к ювенильному состоянию по морфологическим признакам, хотя и усиливает способность к корнеобразованию.
Ювенильное состояние поддерживается специфическим соотношением гормонов. В ряде случаев обработке гиббереллином приводит к образованию ювенильных листьев или к продлению ювенильного состояния.
Переход от ювенильного типа роста к зрелому наиболее хорошо заметен у древесных растений. У бука, например, возможно сосуществование всех фаз развития ювенильности до зрелости. Это свойство древесных растений широко используется в садоводстве, когда для получения ювенильных растений отбирают черенки от основания сеянца, а черенки, взятые из верхних, более зрелых ярусов, укореняют или прививают для получения быстрого зацветающих и плодоносящих растений (Полевой, 1989).
3. Этап зрелости и размножения
На этом этапе происходит формирование генеративных органов и образование плодов. У растений выделяют половое, бесполое и вегетативное размножение. При половом размножении новый организм появляется в результате слияния половых клеток — гамет. Бесполое размножение характерно для споровых растений, у которых чередуются два поколения — бесполое диплоидное и половое гаплоидное. При бесполом размножении новый организм развивается из спор. Вегетативным размножением называют воспроизведение растений из вегетативных частей растения (клубней, луковиц, отводок).
Инициация перехода к цветению осуществляется под действием температуры (яровизация), чередования дня и ночи (фотопериодизм) или эндогенных факторов, обусловленных возрастом растения.
У большинства растений наибольшей чувствительностью к фотопериоду обладают листья, только что закончившие рост. Показано участие в переходе к цветению стимулятора роста гиббереллина.
Цветки, как органы полового размножения, могут быть обоеполыми или раздельнополыми. Они формируются на одних и тех же (однодомность) или на разных (двудомность) растениях. Все факторы внешней среды влияют на гормональный обмен растений, причем факторы, приводящие к сдвигу гормонального баланса в сторону увеличения содержания цитокинина усиливает женскую сексуализацию, а вызывающие увеличение содержания гиббереллина — мужскую. Гормональная система взаимодействует с генетическим аппаратом, что и обусловливает проявление пола растения. Оплодотворение делят на три фазы: а) опыление, б) прорастание пыльцы и рост пыльцевой трубки в тканях пестика, в) собственно оплодотворение, то есть образование зиготы. Плод развивается из завязи цветка и, как правило, содержит семена. Ткани плода формируются из завязи, но нередко в образовании плода участвуют и другие части цветка. Ткани плода закладываются при формировании цветка, причем в ряде случаев плод может развиваться и независимо от семян (партенокарпические, т. е. бессемянные плоды).
Реферат старение населения и цели социальной политики
... экономической сфере старение населения отражается на экономическом росте, накоплениях, инвестициях, потреблении, занятости, пенсионном обеспечении, налоговой политике и передаче накопленных знаний и опыта из поколения в поколение. В социальной сфере старение населения сказывается на ...
Однако обычно цветки без опыления и оплодотворения опадают.
Развитие плодов можно разделить на четыре фазы: 1) формирование завязи до опыления; 2) рост за счет деления клеток сразу после опыления и оплодотворения; 3) рост за счет растяжения клеток; 4) созревание (Полевой,1989).
Цветение — это сложный процесс, критический для жизненного цикла высших растений. Благодаря взаимодействию множества факторов внешней среды и эндогенных регуляторов возникает разветвленная сеть сигнальных путей, контролирующих время перехода к цветению . У растений Arabidopsis известны несколько самостоятельных, хотя и связанных между собой, генетических путей регуляции времени зацветания: автономный, фотопериодический, холодовой. Сигналы от этих путей интегрируются несколькими общими мишенями, которые называют интеграторами времени цветения. Интеграторы времени цветения активируют затем гены идентичности флоральных меристем, и таким образом способствуют переходу растения от вегетативного к репродуктивному развитию.
Для успешного размножения растений чрезвычайно важно точно определить время перехода к цветению. Молекулярно-генетический анализ контроля времени зацветания у растений Arabidopsis выявил интегрированную систему процессов, количественно регулирующих этот переход в развитии растений. Центральная роль в этом переключении принадлежит гену FLOWERING LOCUS C (FLC), который задерживает зацветание, угнетая гены, необходимые для переключения меристемы с вегетативного развития на цветение (Чжицян Янь и др., 2010) .
Ферменты цикла Бенсона-Кальвина могут образовывать мультиферментные комплексы с различным числом ферментов — от двух до восьми, которые различаются по молекулярной массе и функциональным свойствам. Из листьев хлопчатника были выделены свободные мультиферментные комплексы с мол. м. 520 и 240 кД. Мультиферментный комплекс с мол. м. 240 кД был выделен в денатурирующих условиях, как и комплекс из листьев шпината с мол. м. 284 кД. Денатурирующие условия при выделении мультиферментных комплексов были использованы для диссоциации их на мультиферментные комплексы с меньшими величинами молекулярных масс. При сравнительном исследовании онтогенетических изменений функциональной активности мультиферментных комплексов с молекулярной массой 520 и 240 кД обнаружено, что наибольшие величины ферментативных активностей комплексы проявляли в фазах бутонизации и цветения растений (Бабаджанова и др., 2010).
4. Этап старости и отмирания
Этап старости и отмирания включает в себя период от полного прекращения плодоношения до естественной смерти организма. Этот период прогрессирующего ослабления жизнедеятельности.
Продолжительность жизни растений, принадлежащих к разным таксономическим группам и различным жизненным формам, варьирует в очень широких пределах. Это свидетельствует о генетической детерминированности нормы времени жизни каждого вида.
Лекарственные растения, нормализующие деятельность нервной системы, ...
... нервную систему и оказывают другие виды действия. С давних времен цикорий является лекарственным растением. Известный таджикский ученый и врач средневековья Авиценна широко применял препараты цикория при лечении ряда заболеваний, ... рубцеватыми следами на поверхности от отмерших листьев. Закрепляется в почве корневище многочисленными белыми шнуровидными корнями. Листья длиной до 1 м, мечевидно- ...
Старение и смерть — завершающая фаза онтогенеза любого многоклеточного растения, но термин «старение» может быть применим не только целому растению, но и к отдельным его органам — листьям, плодам, частям цветков. Этот вид старения изучен наиболее полно, в то время как механизмы старения целого растения исследованы недостаточно.
Для растения характерно разные типы старения. Однолетние растения отмирают целиком. У многолетних трав ежегодно полностью отмирает надземная часть, а корневая система и другие подземные части остаются жизнеспособными. У многих растений в процессе роста стареют и отмирают ранее образовавшиеся нижние листья. У листопадных деревьев осенью одновременно стареют и опадают все листья.
Процесс старения органа завершается его опадением. Перед опаданием листа или плода в основании черешка или плодоножки образуется отделительный слой, представляющий собой зону, состоящую из клеток, ориентированных под прямыми углами к оси черешка. Размягчаются и частично растворяются клеточные стенки или срединные пластинки клеток отделительной зоны. Этот процесс индуцируется этиленом, вырабатываемом стареющими листьями.
Старение изолированных листьев задерживается при обработке их цитокинином, который стимулирует синтез РНК, белков и аттрагирующую активность клеток. У некоторых растений задержка старения может вызываться также ауксином и гиббереллинами. Абсцизовая кислота и этилен, наоборот, ускоряют процессы старения.
Одной из первых гипотез, объясняющих механизм старения целого растения, была гипотеза немецкого физиолога растений Г. Молиша, основанная на том, что у монокарпических растений — однолетних, двулетних (например, капуста) и некоторых многолетних (агава, бамбук), цветущих лишь один раз, старение наблюдается сразу после цветения и созревание плодов. По мнению Молиша, старение вызывается оттоком большей части питательных веществ к развивающимся репродуктивным органам и отмирание наступает от истощения. Это точка зрения подтверждается тем, что срезание цветков может задержать или даже предотвратить старение целого растения. Против этой гипотезы, однако, говорит тот факт, что удаление мужских цветков у двудомных растений также задерживает старение, хотя значительного оттока ассимилятов к цветкам в данном случае не происходит. Кроме того, у некоторых растений старение индуцируется определенным фотопериодом. Возможно, старение вызывается нарушением соотношения фитогормонов, возникающим как под действием внешних стимулов, так и внутренних факторов.
Основную роль в процессах старения играет функциональная корреляция между корнями и листьями: причиной ослабления роста и наступления старения может быть отставание развития корневой системы. Старение во время формирования плодов определяется угнетением роста корней из-за прекращения поступления в них ассимилятов. Уменьшение активности корней приводит к нарушению водного режима, фотосинтеза, синтеза белков и т. д., что снижает общую жизнедеятельность растений.
Понятие психического старения
... в значительной мере развиваться. Старение – генетически предопределенный этап, и, как и любая стадия жизни человека, сопровождается изменениями физиологии и психологии. Человек не в состоянии ... или урегулировать уменьшение психологической активности. Наступает равновесие между дефицитарными признаками, характеризующими старение, и компенсаторными. Из этого следует, что при позитивном настрое люди ...
У поликарпических растений, цветущих много раз, например у деревьев, по мере увеличения высоты ствола связь между листьями и корнями также ухудшается. Угнетение роста корней, чем бы это ни было вызвано, приводит к уменьшению поступления цитокинина в надземные органы и снижению пролиферативной активности апикальных меристем побега, что и служит причиной старения целого растения (Полевой,1989).
На протяжении онтогенеза монокарпического растения от семени до семени типы метаболизма его тканей неоднократно меняются в соответствии с заложенной в нем генетической программой, реализация которой в значительной степени зависит от действующих внешних факторов и их сочетаний. У двудольных на самой первой стадии онтогенеза прорастание семени под землей и рост первичного корешка, подсемядольного колена и семядолей реализуется только за счет использования запасенных питательных веществ семени. Этот ранний тип углеродного питания принято квалифицировать как гетеротрофный тип питания. Но сразу же после выхода проростка на поверхность по мере развертывания листа начинается формирование фотосинтетического аппарата, и гетеротрофный путь питания листьев в дневной период постепенно заменяется на фотолитоавтотрофный, источником энергии которого является свет, восстановитель — вода и единственный источник углерода СО2. Локализованный в хлоропластах, этот тип метаболизма становится доминирующим в дневное время и достигает своего максимума приблизительно ко времени полного развертывания листа. Конечный этап старения листа — «дряхление» − это необратимый процесс, завершающийся отмиранием листа. На протяжении всего жизненного цикла монокарпического растения происходит последовательная утеря старых и образование новых молодых листьев. У однолетних монокарпиков старение листа, включая и его гибель, — естественный физиологический процесс, квалифицируемый как нормальная стадия развития. На всем ee протяжении происходит снижение фотоавтотрофной функции, причем листья-акцепторы превращаются в доноров метаболитов. При этом наблюдается кардинальная перестройка метаболизма, состоящая в распаде одних биохимических компонентов и замене их на другие и сопровождающаяся оттоком их в другие органы-акцепторы. Если есть «запрос» со стороны акцепторов, углерод, азот и другие элементы, содержащиеся в продуктах частичного распада метаболитов, используются на поддержание роста корней и новых растущих молодых листьев. В период старения жизнедеятельность листового аппарата растения может продолжаться до полного истощения ресурсов листа-донора. Отток продуктов распада из стареющих листьев может направляться в нижние, верхние органы и в обоих направлениях в зависимости от силы «запросов» акцепторов. Продолжительность и эффективность работы листового аппарата зависят от условий выращивания и, насколько позволяет продолжительность вегетационного периода, связаны с продуктивностью фотосинтеза и всего растения. Старение листа, как и специфика всего онтогенеза, управляется как на уровне метаболической регуляции, так и на уровне биосинтезов с участием высоко эффективных сигнальных реакций, идущих как от метаболитов так и от фитогормонов (Романова и др., 2011).
Генетические основы онтогенеза
... типы онтогенеза; цитогенетические основы дифференцировки в онтогенезе; явление тотипотентности соматических клеток; дифференциальную активность генов в онтогенезе. 1. Генетические основы онтогенеза Онтогенезом называется индивидуальное ... за счет обмена метаболитами между органами и частями растений. В онтогенезе семенных растений выделяют следующие периоды: 1. предзародышевый (преэмбриональный) – ...
Заключение
Таким образом, в ходе онтогенеза растительные организмы проходят ряд этапов: эмбриональный, ювенильный, зрелости и размножения, старости и отмирания. Каждый из этих этапов в свою очередь включает в себя несколько последовательных фаз роста и развития.
В начале своего развития зародыши зависят от фитогормонов, поступающих из других тканей, прежде всего из эндосперма. Затем они сами начинают синтезировать фитогормоны, а при формировании семян ауксин выделяется из них в окружающие ткани, активируя рост околоплодника
В этап молодости осуществляется прорастание семян и формирование вегетативных органов. Растения на этом этапе не способны к половому размножению.
В этап зрелости и размножения происходит формирование генеративных органов и образование плодов.
Старение имеет большое биологическое значение. Оно является одним из способов адаптации растений к неблагоприятным условиям внешней среды. Кроме того, старение способствует более быстрой эволюции, так как ускоряет смену поколений.
Список литературы
[Электронный ресурс]//URL: https://psystars.ru/referat/etapyi-ontogeneza-ovoschnyih-rasteniy/
1. Бабаджанова М. А., Мирзорахимов А. К., Бабаджанова М. П., Эсаналиева Ш. А. Онтогенетическая зависимость образования различных мультиферментных комплексов цикла Бенсона-Кальвина в листьях хлопчатника // Физиология растений.2010. Т. 57, № 2, С.186-191.
— Мейчик Н. Р., Смирнова А. В., Матвеева Н. П., Николаева Ю. И., Ермаков И. П. Изменение состава ионогенных групп оболочки пыльцевого зерна лилии при активации прорастания // Физиология растений. 2009. Т. 56, № 2, С.232-240.
— Полевой В. В. Физиология растений: Учеб. Для биол. спец. вузов. — М.: Высш. шк., 1989. — 464 с.
— Романова А. К., Семенова Г. А., Новичкова Н. С., Игнатьев А. Р., Мудрик В. А., Иванов Б. Н. Физиолого-биохимические и флуоресцентные показатели старения листьев сахарной свеклы в вегетативной фазе роста // Физиология растений. 2011. Т. 58, № 2, С.221-233.
— Чжицян Янь, Давэй Лян, Хен Лю, Гочан Чжэнь FLC: ключевой регулятор времени зацветания у Arabidopsis // Физиология растений. 2010. Т. 57, № 2, С.177-185.